Fotoatlas katedry biologie FPE ZČU
Hledat:
-
 Anarthophyllum strigulipetalum SorarúFabaceae Tvorí asi 50 cm vysoké a více než metr velké ke?íky s nápadnými kv?ty. Kvete brzy na ja?e. Druh je rozší?ený a p?vodní v Patagonii v Jižní Americe na skalnatém podkladu až do 900 m n.m. © Michal Mergl, 2015
Anarthophyllum strigulipetalum SorarúFabaceae Tvorí asi 50 cm vysoké a více než metr velké ke?íky s nápadnými kv?ty. Kvete brzy na ja?e. Druh je rozší?ený a p?vodní v Patagonii v Jižní Americe na skalnatém podkladu až do 900 m n.m. © Michal Mergl, 2015 -
 Anemochorie - Eryngium campestreZvláštní typ anemochorie mají tzv. stepní běžci (např. máčka ladní). Současně se zráním plodů usychá celá rostlina, která se díky činnosti větru pohybuje po zemi ("kutálí") a semena se tak uvolňují na velké ploše. © T.Č. Kučera 2006
Anemochorie - Eryngium campestreZvláštní typ anemochorie mají tzv. stepní běžci (např. máčka ladní). Současně se zráním plodů usychá celá rostlina, která se díky činnosti větru pohybuje po zemi ("kutálí") a semena se tak uvolňují na velké ploše. © T.Č. Kučera 2006 -
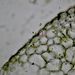
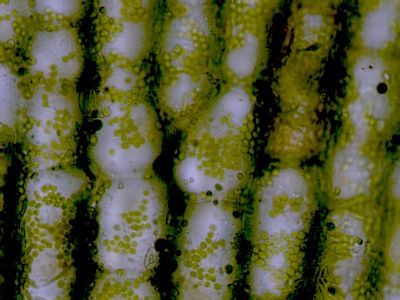
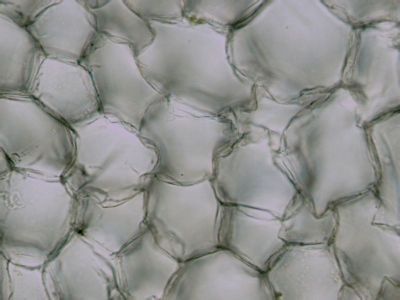
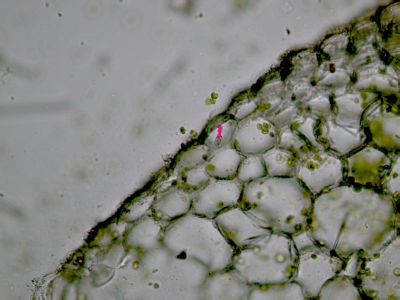
 Asimilační pletivo - houbový parenchymHoubový parenchym je součástí asimilačního pletiva v listu - mezofylu. Je tvořen nepravidelně větvenými buňkami s velkými intercelulárami. Připomíná plektenchym hub - proto se nazývá houbový. © T.Č. Kučera 2006
Asimilační pletivo - houbový parenchymHoubový parenchym je součástí asimilačního pletiva v listu - mezofylu. Je tvořen nepravidelně větvenými buňkami s velkými intercelulárami. Připomíná plektenchym hub - proto se nazývá houbový. © T.Č. Kučera 2006 -
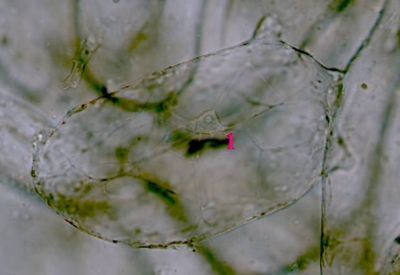
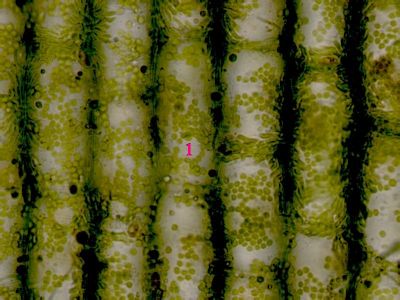
 Asimilační pletivo Elodea canadensisParenchymatické asimilační pletivo z listu vodního moru kanadského. Buňky obsahují zelené plastidy - chloroplasty - kulovitého tvaru. Okrouhlé struktury (1), které jsou chloroplasty obklopeny, jsou buněčná jádra. © T.Č. Kučera 2006
Asimilační pletivo Elodea canadensisParenchymatické asimilační pletivo z listu vodního moru kanadského. Buňky obsahují zelené plastidy - chloroplasty - kulovitého tvaru. Okrouhlé struktury (1), které jsou chloroplasty obklopeny, jsou buněčná jádra. © T.Č. Kučera 2006 -
 Buňky dužniny šípkuV parenchymatických buňkách šípku (Rosa sp.) jsou patrné podlouhlé oranžové chromoplasty (1) a drúzy krystalů šťavelanu vápenatého. © T.Č. Kučera 2006
Buňky dužniny šípkuV parenchymatických buňkách šípku (Rosa sp.) jsou patrné podlouhlé oranžové chromoplasty (1) a drúzy krystalů šťavelanu vápenatého. © T.Č. Kučera 2006 -
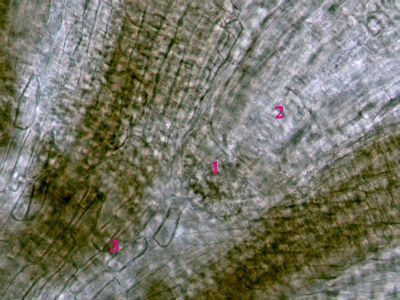
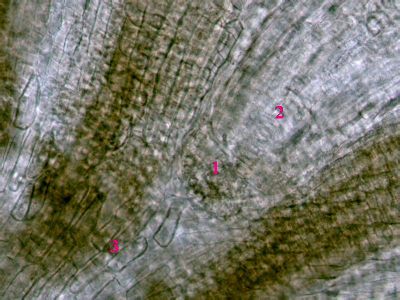
 Dřevní parenchymSoučástí cévních svazků a vodivých pletiv jsou i složky, které se nepodílejí bezprostředním způsobem na vedení roztoků. jde zejméan o tzv. dřevní a lýkový parenchym, které probíhají paralelně s vodivými drahami. Tzv. radiální (paprskový) parenchym (1) naproti tomu prochází ve směru kolmém na vodivé dráhy. Dřevo Pinus sylvestris. © T.Č. Kučera 2006
Dřevní parenchymSoučástí cévních svazků a vodivých pletiv jsou i složky, které se nepodílejí bezprostředním způsobem na vedení roztoků. jde zejméan o tzv. dřevní a lýkový parenchym, které probíhají paralelně s vodivými drahami. Tzv. radiální (paprskový) parenchym (1) naproti tomu prochází ve směru kolmém na vodivé dráhy. Dřevo Pinus sylvestris. © T.Č. Kučera 2006 -
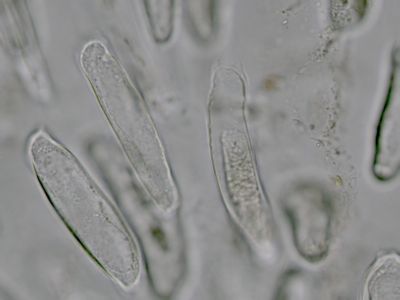
 Dvůrkaté tečkyDvůrkaté tečky přestavují ztenčeniny buněčné stěny, jimiž mohou komunikovat sousední buňky. V tomto případě dvě buňky tracheidy, které mají zachované příčné přepažky. Vlastní otvor ve stěně (porus) je zobrazen jako menší tečka, dvůrek kolem je část ztlustlé stěny. Řez dřevem borovice lesní (Pinus sylvetris). © T.Č. Kučera 2006
Dvůrkaté tečkyDvůrkaté tečky přestavují ztenčeniny buněčné stěny, jimiž mohou komunikovat sousední buňky. V tomto případě dvě buňky tracheidy, které mají zachované příčné přepažky. Vlastní otvor ve stěně (porus) je zobrazen jako menší tečka, dvůrek kolem je část ztlustlé stěny. Řez dřevem borovice lesní (Pinus sylvetris). © T.Č. Kučera 2006 -
 EntomogamieOpylení hmyzem představuje (ještě vedle anemogamie) hlavní způsob opylení u krytosemenných rostlin. Vznik tohoto jevu je vysvětlován tzv. koevolucí, tj. současným vývojem dvou skupin organismů, v tomto případě entomogamních rostlin a hmyzu živícího se nektarem, příp. pylem. © T.Č. Kučera 2006
EntomogamieOpylení hmyzem představuje (ještě vedle anemogamie) hlavní způsob opylení u krytosemenných rostlin. Vznik tohoto jevu je vysvětlován tzv. koevolucí, tj. současným vývojem dvou skupin organismů, v tomto případě entomogamních rostlin a hmyzu živícího se nektarem, příp. pylem. © T.Č. Kučera 2006 -
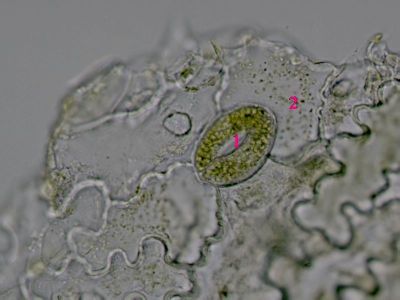
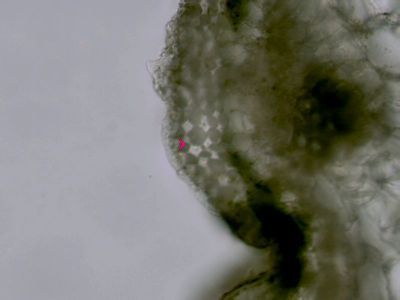
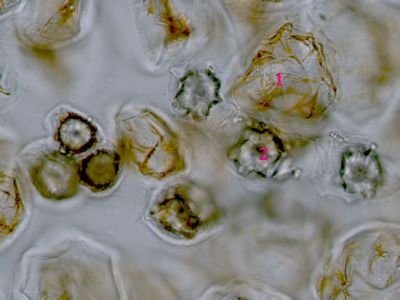
 Ergastické produkty - šťavelan vápenatýRafidy (1) a dipyramida (2) šťavelanu vápenatého ze šťávy begónie (Begonia sp.). © T.Č. Kučera 2006
Ergastické produkty - šťavelan vápenatýRafidy (1) a dipyramida (2) šťavelanu vápenatého ze šťávy begónie (Begonia sp.). © T.Č. Kučera 2006 -
 GutaceGutace umožňuje rostlině zbavovat se vody v době, kdy vysoká relativní vzdušná vlhkost zabraňuje transpiraci. Tento jev probíhá metamorfovanými průduchy - hydatodami - které ztratily schopnost uzvírání. © T.Č. Kučera 2006
GutaceGutace umožňuje rostlině zbavovat se vody v době, kdy vysoká relativní vzdušná vlhkost zabraňuje transpiraci. Tento jev probíhá metamorfovanými průduchy - hydatodami - které ztratily schopnost uzvírání. © T.Č. Kučera 2006 -
 Hloh (Crataegus)Na záběru jsou patrné květy rostliny čel. Rosaceae: aktinomorfní, heterochlamydní, pětičetné, s mnoha tyčinkami. Zašpičatělé krátké větévky jsou tzv. kolce, trny stonkového původu. Květy jsou entomogamní - zde je opyluje zlatohlávek. © T.Č. Kučera 2006
Hloh (Crataegus)Na záběru jsou patrné květy rostliny čel. Rosaceae: aktinomorfní, heterochlamydní, pětičetné, s mnoha tyčinkami. Zašpičatělé krátké větévky jsou tzv. kolce, trny stonkového původu. Květy jsou entomogamní - zde je opyluje zlatohlávek. © T.Č. Kučera 2006 -
 Hojivá pletiva - závalZával neboli kalus představuje typ nediferencovaných pletiv, která postupně zacelují ránu. Následuje proces diferenciace a funkční specializace. © T.Č. Kučera 2006
Hojivá pletiva - závalZával neboli kalus představuje typ nediferencovaných pletiv, která postupně zacelují ránu. Následuje proces diferenciace a funkční specializace. © T.Č. Kučera 2006 -
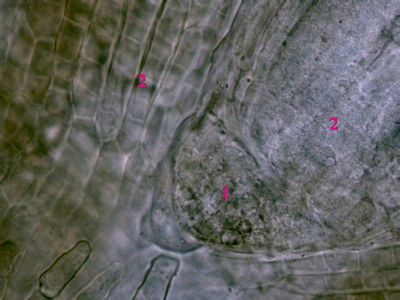
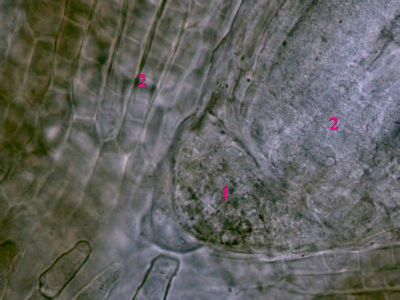
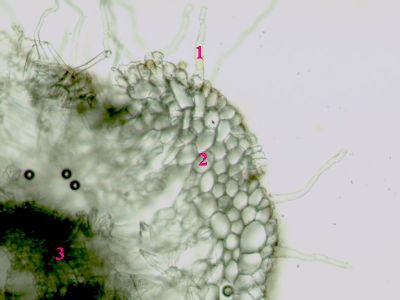
 Iniciála v kořeni Ranunculus repensIniciála (1) představuje typ protomeristému - tedy nejpůvodnějšího meristému v rostlině. Je lokalizována ve vzrostném vrcholu, jde tedy o apikální meristém. Z ní vycházejí primární meristémy (2), které produkují primární vodivá a krycí pletiva. © T.Č. Kučera 2006
Iniciála v kořeni Ranunculus repensIniciála (1) představuje typ protomeristému - tedy nejpůvodnějšího meristému v rostlině. Je lokalizována ve vzrostném vrcholu, jde tedy o apikální meristém. Z ní vycházejí primární meristémy (2), které produkují primární vodivá a krycí pletiva. © T.Č. Kučera 2006 -
 Iniciála v kořeni Ranunculus repens - lokalizaceIniciála (1) je umístěna ve střední části kořenové špičky. Směrem vzhůru jsou patrná primární vodivá pletiva (2), dolů ke špičce kořene následuje střední sloupek (columela) se statolitickým škrobem (3) a kalyptrogen - meristém produkující kořenovou čepičku. © T.Č. Kučera 2006
Iniciála v kořeni Ranunculus repens - lokalizaceIniciála (1) je umístěna ve střední části kořenové špičky. Směrem vzhůru jsou patrná primární vodivá pletiva (2), dolů ke špičce kořene následuje střední sloupek (columela) se statolitickým škrobem (3) a kalyptrogen - meristém produkující kořenovou čepičku. © T.Č. Kučera 2006 -
 Kalyptra (kořenové čepička)Detail buňek kořenové čepičky, které jsou vyplněny slizem. © T.Č. Kučera 2006
Kalyptra (kořenové čepička)Detail buňek kořenové čepičky, které jsou vyplněny slizem. © T.Č. Kučera 2006 -
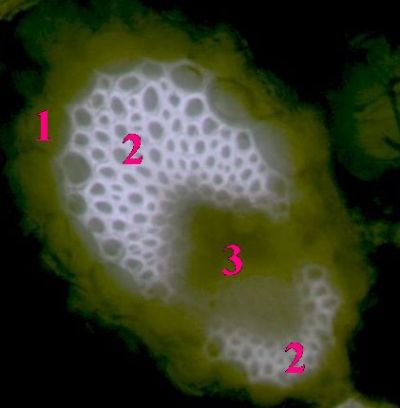
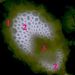 Kolaterální cévní svazekNa obrázku je patrná parenchymatická pochva cévního svazku (1), sklerenchymatická pletiva (2) a pak vlastní vodivá pletiva (3)- © T.Č. Kučera 2006
Kolaterální cévní svazekNa obrázku je patrná parenchymatická pochva cévního svazku (1), sklerenchymatická pletiva (2) a pak vlastní vodivá pletiva (3)- © T.Č. Kučera 2006 -
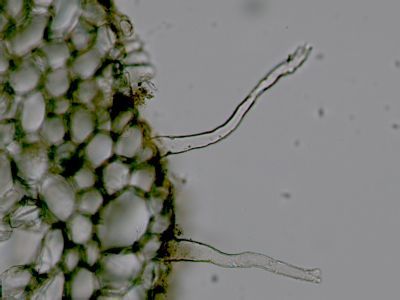
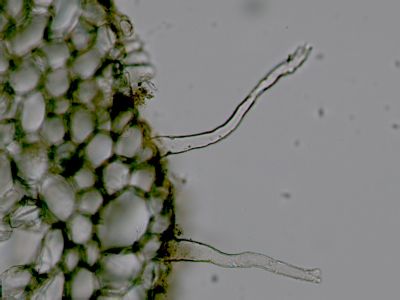
 Kořen - detail báze kořenového vláskuKořenový vlásek je derivátem rhizodermis, podobně jako trichomy u epidermis. Na rozdíl od trichomů jsou kořenové vlásky vždy jednobuněčné a živé. © T.Č. Kučera 2006
Kořen - detail báze kořenového vláskuKořenový vlásek je derivátem rhizodermis, podobně jako trichomy u epidermis. Na rozdíl od trichomů jsou kořenové vlásky vždy jednobuněčné a živé. © T.Č. Kučera 2006 -
 Kořen - roztlak kořené špičky (detail)Detailní pohled na buňky kořenové čepičky. © T.Č. Kučera 2006
Kořen - roztlak kořené špičky (detail)Detailní pohled na buňky kořenové čepičky. © T.Č. Kučera 2006 -
 Kořen - roztlak kořenové špičkyV záběru jsou nápadné parenchymtické buňky, radiálně se rozbíhající od kořenové špičky. Jde o tlakem odlouplé buňky kalyptry (kořenové čepičky). © T.Č. Kučera 2006
Kořen - roztlak kořenové špičkyV záběru jsou nápadné parenchymtické buňky, radiálně se rozbíhající od kořenové špičky. Jde o tlakem odlouplé buňky kalyptry (kořenové čepičky). © T.Č. Kučera 2006 -
 Kořen Ranunculus repens - kořenové vlášeníV absorpční zóně kořene je poměrně hustý porost kořenových vlásků, které několikanásobně zvyšují aktivní opvrch kořene. © T.Č. Kučera 2006
Kořen Ranunculus repens - kořenové vlášeníV absorpční zóně kořene je poměrně hustý porost kořenových vlásků, které několikanásobně zvyšují aktivní opvrch kořene. © T.Č. Kučera 2006 -
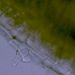 Kořen Ranunculus repens - odlupující se buňka kořenové čepičkyKořenová čepička (kalyptra) vzniká v meristému kalyptrogenu. Je tvořena parenchymatickými buňkami vyplněnými slizem, který usnadňuje pronikání kořene do půdy. © T.Č. Kučera 2006
Kořen Ranunculus repens - odlupující se buňka kořenové čepičkyKořenová čepička (kalyptra) vzniká v meristému kalyptrogenu. Je tvořena parenchymatickými buňkami vyplněnými slizem, který usnadňuje pronikání kořene do půdy. © T.Č. Kučera 2006 -
 Kořenová špička Ranunculus repensDetail kořenové špičky s parenchymatickými buňkami kalyptry. © T.Č. Kučera 2006
Kořenová špička Ranunculus repensDetail kořenové špičky s parenchymatickými buňkami kalyptry. © T.Č. Kučera 2006 -
 Kořeny - chůdovité kořeny Zea maysKukuřice má, podobně jako některé tropické stromy, tzv. chůdovité kořeny. Jde o soubor adventivních kořenů, které vyrůstají nad zemí, "rozepírají" se do stran a zvyšují tak stabilitu stonku. © T.Č. Kučera 2006
Kořeny - chůdovité kořeny Zea maysKukuřice má, podobně jako některé tropické stromy, tzv. chůdovité kořeny. Jde o soubor adventivních kořenů, které vyrůstají nad zemí, "rozepírají" se do stran a zvyšují tak stabilitu stonku. © T.Č. Kučera 2006 -
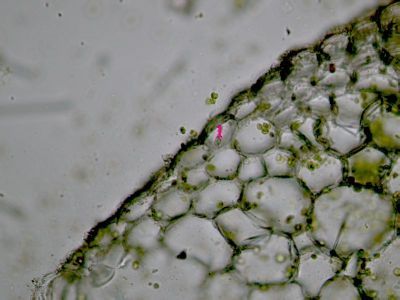
 Krycí pletiva primární - epidermis Polypodium vulgareEpidermis vývojově primitivních cévnatých rostlin obsahuje chloroplasty a má tedy asimilační funkci. Na obrázku je epidermis osladiče obecného s průduchem (1). Buňky vlastní epidermis mají laločnaté perikliny a jsou v nich patrné drobné chloroplasty. © T.Č. Kučera 2006
Krycí pletiva primární - epidermis Polypodium vulgareEpidermis vývojově primitivních cévnatých rostlin obsahuje chloroplasty a má tedy asimilační funkci. Na obrázku je epidermis osladiče obecného s průduchem (1). Buňky vlastní epidermis mají laločnaté perikliny a jsou v nich patrné drobné chloroplasty. © T.Č. Kučera 2006 -
 Krycí pletiva sekundární - borka Robinia pseudacaciaNahromaděním vrstev korku (felem) vzniká borka (rhytidoma), která má funkci sekundárního krycícho pletiva. Vzniká činností sekundárního meristému felogenu. Je-li činnost felogenu kontinuální, vzniká hladká borka jako např. u buku; v případě nepravidleností v činnosti felogenu se vytváří borka zbrázděná - foto povrchu kmene trnovníku akátu. © T.Č. Kučera 2006
Krycí pletiva sekundární - borka Robinia pseudacaciaNahromaděním vrstev korku (felem) vzniká borka (rhytidoma), která má funkci sekundárního krycícho pletiva. Vzniká činností sekundárního meristému felogenu. Je-li činnost felogenu kontinuální, vzniká hladká borka jako např. u buku; v případě nepravidleností v činnosti felogenu se vytváří borka zbrázděná - foto povrchu kmene trnovníku akátu. © T.Č. Kučera 2006 -
 Květ Nonnea pullaKvět piply osmahlé patří mezi vůbec nejtmavší květy v naší flóře. Květy jsou aktinomorfní, heterochlamydní a pětičetné. Rostlina patří mezi brutnákovité (Boraginaceae), jejím plodem jsou tedy tvrdky. © T.Č. Kučera 2006
Květ Nonnea pullaKvět piply osmahlé patří mezi vůbec nejtmavší květy v naší flóře. Květy jsou aktinomorfní, heterochlamydní a pětičetné. Rostlina patří mezi brutnákovité (Boraginaceae), jejím plodem jsou tedy tvrdky. © T.Č. Kučera 2006 -
 Květ Robinia pseudacaciaKvěty trnovníku akátu jsou jako u všech bobovitých (Fabaceae) zygomorfní, heterochlamydní, stavěné podle čísla 5. Petaly jsou označovány jako pavéza (1), křídla (2) a člunek (3). V něm jsou skryté reprodukční orgány - jednoplodolistový pestík a 10 tyčinek. © T.Č. Kučera 2006
Květ Robinia pseudacaciaKvěty trnovníku akátu jsou jako u všech bobovitých (Fabaceae) zygomorfní, heterochlamydní, stavěné podle čísla 5. Petaly jsou označovány jako pavéza (1), křídla (2) a člunek (3). V něm jsou skryté reprodukční orgány - jednoplodolistový pestík a 10 tyčinek. © T.Č. Kučera 2006 -
 Květ Adonis vernalisPryskyřníkovité rostliny (Ranunculaceae) mají fylogeneticky primitivní stavbu květů. Gynaeceum je apokarpní, vícepestíkové. Tyčinek je mnoho. Všechny květní části vyrůstají na vystouplém květním lůžku ve šroubovici. © T.Č. Kučera 2006
Květ Adonis vernalisPryskyřníkovité rostliny (Ranunculaceae) mají fylogeneticky primitivní stavbu květů. Gynaeceum je apokarpní, vícepestíkové. Tyčinek je mnoho. Všechny květní části vyrůstají na vystouplém květním lůžku ve šroubovici. © T.Č. Kučera 2006 -
 Květ Aquilegia vulgarisOrlíček obecný (čel.Ranunculaceae) má pro svoji čeleď netypickou stavbu květů. Nápadné lístky, vyčnívající před květ (1) jsou sepaly, zatímco hlavy "sedících orlů" (2) jsou tvořeny petaly. © T.Č. Kučera 2006
Květ Aquilegia vulgarisOrlíček obecný (čel.Ranunculaceae) má pro svoji čeleď netypickou stavbu květů. Nápadné lístky, vyčnívající před květ (1) jsou sepaly, zatímco hlavy "sedících orlů" (2) jsou tvořeny petaly. © T.Č. Kučera 2006 -
 Květ Campanula persicifoliaZvonkovité Campanulaceae mají aktinomorfní heterochlamydní pětičetné květy. Na obrázku je patrné rovněž mléčení po odlomení horní části stonku. © T.Č. Kučera 2006
Květ Campanula persicifoliaZvonkovité Campanulaceae mají aktinomorfní heterochlamydní pětičetné květy. Na obrázku je patrné rovněž mléčení po odlomení horní části stonku. © T.Č. Kučera 2006 -
 Květ Canna indicaKvěty dosny indické jsou asymetrické. © T.Č. Kučera 2006
Květ Canna indicaKvěty dosny indické jsou asymetrické. © T.Č. Kučera 2006 -
 Květ Euonymus europaeusKvěty brslenu evropského jsou čtyřčetné, stejně jako plody - tobolky, nazývané lidově "kvadrátky". © T.Č. Kučera 2006
Květ Euonymus europaeusKvěty brslenu evropského jsou čtyřčetné, stejně jako plody - tobolky, nazývané lidově "kvadrátky". © T.Č. Kučera 2006 -
 Květ Chelidonium majusKvět vlaštovičníku většího je aktinomorfní heterochlamydní (s prchavým kalichem) a čtyřčetný. Plodem je tobolka. Makovité (Papaveraceae) jsou význačné přítomností mléčnic. © T.Č. Kučera 2006
Květ Chelidonium majusKvět vlaštovičníku většího je aktinomorfní heterochlamydní (s prchavým kalichem) a čtyřčetný. Plodem je tobolka. Makovité (Papaveraceae) jsou význačné přítomností mléčnic. © T.Č. Kučera 2006 -
 Květ Linum austriacumKvět lnu je aktinomrfní, heterochlamydní, stavěný podle čísla 5. To se odráží v počtu tyčinek i pačnělek (stylodií). © T.Č. Kučera 2006
Květ Linum austriacumKvět lnu je aktinomrfní, heterochlamydní, stavěný podle čísla 5. To se odráží v počtu tyčinek i pačnělek (stylodií). © T.Č. Kučera 2006 -
 Květ PaeoniaV květech pivoňky jsou přítomny některé vývojově primitivní znaky. Na fotografii po odkvětu je patrný plynulý přechod mezi listeny (1) a kališními lístky (2), s výskytem několika přechodných typů. To vedlo k dodnes akceptované myšlence listenového původu sepalů. © T.Č. Kučera 2006
Květ PaeoniaV květech pivoňky jsou přítomny některé vývojově primitivní znaky. Na fotografii po odkvětu je patrný plynulý přechod mezi listeny (1) a kališními lístky (2), s výskytem několika přechodných typů. To vedlo k dodnes akceptované myšlence listenového původu sepalů. © T.Č. Kučera 2006 -
 Květ snědku (Ornithogallum sp.)Snědek patří mezi rostliny liliovité (Liliaceae). Květ je homochlamydní, tvořený dvěma kruhy tepalů po třech. I tyčinky a pestík jsou vystaveny podle čísla 3. © T.Č. Kučera 2006
Květ snědku (Ornithogallum sp.)Snědek patří mezi rostliny liliovité (Liliaceae). Květ je homochlamydní, tvořený dvěma kruhy tepalů po třech. I tyčinky a pestík jsou vystaveny podle čísla 3. © T.Č. Kučera 2006 -
 Květenství Morus sp.Morušovník má jednoduchá hroznovitá květenství, která dávají vznik plodenstvím - moruším. Ty jsou sice vnějšími znaky podobné např. ostružinám, ale v případě ostružin jde o souplodí, tedy útvar vzniklý z jednoho květu s více apokarpními pestíky. © T.Č. Kučera 2006
Květenství Morus sp.Morušovník má jednoduchá hroznovitá květenství, která dávají vznik plodenstvím - moruším. Ty jsou sice vnějšími znaky podobné např. ostružinám, ale v případě ostružin jde o souplodí, tedy útvar vzniklý z jednoho květu s více apokarpními pestíky. © T.Č. Kučera 2006 -
 Květenství Sambucus nigraKvětenstvím bezu černého je vrcholík mnohoramenný - pleiochasium. Jde o výchozí typ vrcholičnatých květenství, kdy hlavní vřeteno květenství přerůstá více postranních větví. Plodem bezu je peckovice. © T.Č. Kučera 2006
Květenství Sambucus nigraKvětenstvím bezu černého je vrcholík mnohoramenný - pleiochasium. Jde o výchozí typ vrcholičnatých květenství, kdy hlavní vřeteno květenství přerůstá více postranních větví. Plodem bezu je peckovice. © T.Č. Kučera 2006 -
 Květenství Viburnum lantanaKvětenstvím kaliny tušalaje je, podobně jako u bezu černého, vrcholík mnohoramenný (pleiochasium). © T.Č. Kučera 2006
Květenství Viburnum lantanaKvětenstvím kaliny tušalaje je, podobně jako u bezu černého, vrcholík mnohoramenný (pleiochasium). © T.Č. Kučera 2006 -
 Květenství Zea maysKukuřice má dva různé typy květenství: samičí jsou palice vyrůstající v úžlabí listů, samčí je vrcholová lata. © T.Č. Kučera 2006
Květenství Zea maysKukuřice má dva různé typy květenství: samičí jsou palice vyrůstající v úžlabí listů, samčí je vrcholová lata. © T.Č. Kučera 2006 -
 Květy Crataegus sp.Květy hlohu jsou aktinomorfní, heterochlamydní, K5, C5, A mnoho, G 3-5 (apokarpní), plodem je malvice. © T.Č. Kučera 2006
Květy Crataegus sp.Květy hlohu jsou aktinomorfní, heterochlamydní, K5, C5, A mnoho, G 3-5 (apokarpní), plodem je malvice. © T.Č. Kučera 2006 -
 Květy Lamium albumKvěty hluchavky bílé jsou zygomorfní, heterochlamydní, synsepalní a synpetalní. Tyčinky jsou čtyři a jsou dvoumocné. Květenstvím hluchavky je lichopřeslen, plodem tvrdka. © T.Č. Kučera 2006
Květy Lamium albumKvěty hluchavky bílé jsou zygomorfní, heterochlamydní, synsepalní a synpetalní. Tyčinky jsou čtyři a jsou dvoumocné. Květenstvím hluchavky je lichopřeslen, plodem tvrdka. © T.Č. Kučera 2006 -
 Květy a květenství Primula verisKvěty prvosenky jarní jsou aktinomrofní, heterochlamydní, synsepalní a synpetalní. Determinačním znakem oproti Primula elatior jsou nafouklé kalichy. Květy jsou uspořádány do okolíku, na jehož bázi vyrůstají obaly listenového původu. © T.Č. Kučera 2006
Květy a květenství Primula verisKvěty prvosenky jarní jsou aktinomrofní, heterochlamydní, synsepalní a synpetalní. Determinačním znakem oproti Primula elatior jsou nafouklé kalichy. Květy jsou uspořádány do okolíku, na jehož bázi vyrůstají obaly listenového původu. © T.Č. Kučera 2006 -
 Květy a květenství Primula veris - detailKvěty prvosenky jarní jsou aktinomrofní, heterochlamydní, synsepalní a synpetalní. Determinačním znakem oproti Primula elatior jsou nafouklé kalichy. Květy jsou uspořádány do okolíku, na jehož bázi vyrůstají obaly listenového původu. © T.Č. Kučera 2006
Květy a květenství Primula veris - detailKvěty prvosenky jarní jsou aktinomrofní, heterochlamydní, synsepalní a synpetalní. Determinačním znakem oproti Primula elatior jsou nafouklé kalichy. Květy jsou uspořádány do okolíku, na jehož bázi vyrůstají obaly listenového původu. © T.Č. Kučera 2006 -
 Lata tvořená klásky (Arrhenatherum elatius)Květenstvím všech trav (čel. Poaceae) je klásek. Jde o značně redukované květenství, tvořené jedním až třema květy. Zachovány jsou obvykle listence (plevy), listen (plucha, někdy s osinou), část vnějšího kruhu okvětí (pluška) a vniřní kruh okvětí (plenky). Tyčinky jsou tři s vrtivým prašníkem (vztah k anemogamii) a pestík má zpeřenou dvouklanou bliznu. © T.Č. Kučera 2006
Lata tvořená klásky (Arrhenatherum elatius)Květenstvím všech trav (čel. Poaceae) je klásek. Jde o značně redukované květenství, tvořené jedním až třema květy. Zachovány jsou obvykle listence (plevy), listen (plucha, někdy s osinou), část vnějšího kruhu okvětí (pluška) a vniřní kruh okvětí (plenky). Tyčinky jsou tři s vrtivým prašníkem (vztah k anemogamii) a pestík má zpeřenou dvouklanou bliznu. © T.Č. Kučera 2006 -
 List - jednoduchý list Carpinus betulusListy habru obecného mají jednoduchou čepel, zpeřenou žilnatinu a dvakrát pilovitý okraj. © T.Č. Kučera 2006
List - jednoduchý list Carpinus betulusListy habru obecného mají jednoduchou čepel, zpeřenou žilnatinu a dvakrát pilovitý okraj. © T.Č. Kučera 2006 -
 List - lichozpeřený list Fraxinus excelsiorLichozpeřený složený list jasanu ztepilého. Lístky tvoří jařma (páry) přisedající ke střednímu žebru. © T.Č. Kučera 2006
List - lichozpeřený list Fraxinus excelsiorLichozpeřený složený list jasanu ztepilého. Lístky tvoří jařma (páry) přisedající ke střednímu žebru. © T.Č. Kučera 2006 -
 List - lichozpeřený list Robinia pseudacaciaListy trnovníku akátu jsou složené lichozpeřené a jejich lístky mají řapíčky. Palisty na bázi jsou přeměněny v trny. © T.Č. Kučera 2006
List - lichozpeřený list Robinia pseudacaciaListy trnovníku akátu jsou složené lichozpeřené a jejich lístky mají řapíčky. Palisty na bázi jsou přeměněny v trny. © T.Č. Kučera 2006 -
 List - přenodílný list Crataegus sp.Jednoduchý členěný list tohoto hlohu má čepel peřenodílnou, tj. výkrojky zasahují do cca 2/3 šířky listu (= laločný) a žilnatina je tvořena hlavní žilkou, z níž vybíhají žilky postranní (= zpeřená žilnatina). © T.Č. Kučera 2006
List - přenodílný list Crataegus sp.Jednoduchý členěný list tohoto hlohu má čepel peřenodílnou, tj. výkrojky zasahují do cca 2/3 šířky listu (= laločný) a žilnatina je tvořena hlavní žilkou, z níž vybíhají žilky postranní (= zpeřená žilnatina). © T.Č. Kučera 2006 -
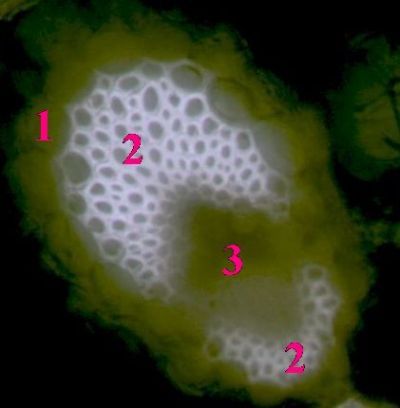
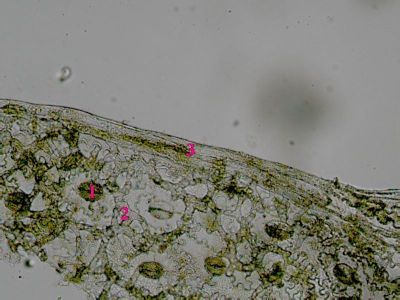
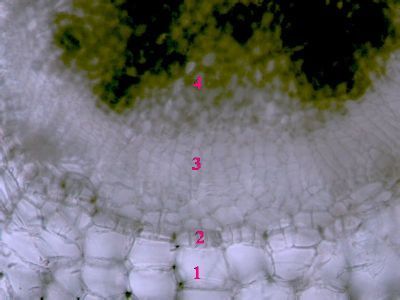
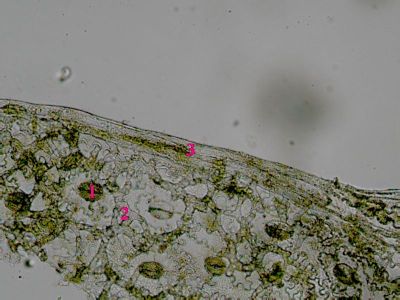
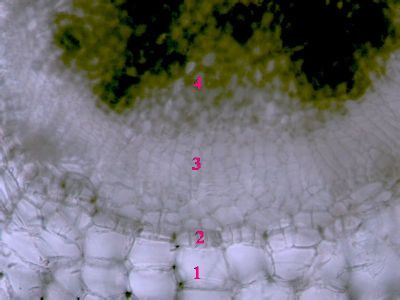
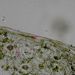 List Polypodium vulgarePohled na list osladiče s patrnou epidermis s průduchy (1) a o něco nížeji položenou vrstvu houbovitého parnechymu (2). Na okraji listu jsou buňky prosenchymatického typu se šikmými příčnými přepážkami (3). © T.Č. Kučera 2006
List Polypodium vulgarePohled na list osladiče s patrnou epidermis s průduchy (1) a o něco nížeji položenou vrstvu houbovitého parnechymu (2). Na okraji listu jsou buňky prosenchymatického typu se šikmými příčnými přepážkami (3). © T.Č. Kučera 2006 -
 List Quercus petraeaDub zimní má peřenolaločné listy. Báze listové čepele je sbíhavá, což je jeden z determinačních znaků oproti dubu letnímu. Zřejmé je střídavé postavení listu a zpeřená žilnatina. © T.Č. Kučera 2006
List Quercus petraeaDub zimní má peřenolaločné listy. Báze listové čepele je sbíhavá, což je jeden z determinačních znaků oproti dubu letnímu. Zřejmé je střídavé postavení listu a zpeřená žilnatina. © T.Č. Kučera 2006 -
 List jednoděložné rostliny - Zea maysKukuřice má podobnou stavbu listů jako ostatní trávy: listy nemají řapík, ale ke stonku přirůstají v kolénku (nodu) a jako listová pochva (1) objímají stonek. Na ni navazuje čepel (2). © T.Č. Kučera 2006
List jednoděložné rostliny - Zea maysKukuřice má podobnou stavbu listů jako ostatní trávy: listy nemají řapík, ale ke stonku přirůstají v kolénku (nodu) a jako listová pochva (1) objímají stonek. Na ni navazuje čepel (2). © T.Č. Kučera 2006 -
 List, plod a větévka Castanea sativaKaštanovník setý, patřící do čel. Fagaceae má na rozdíl od jírovce jednoduché listy s pilovitým okrajem. Plodem je nažka v číšce (u jírovce tobolka). © T.Č. Kučera 2006
List, plod a větévka Castanea sativaKaštanovník setý, patřící do čel. Fagaceae má na rozdíl od jírovce jednoduché listy s pilovitým okrajem. Plodem je nažka v číšce (u jírovce tobolka). © T.Č. Kučera 2006 -
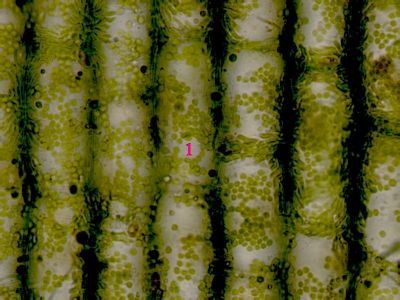
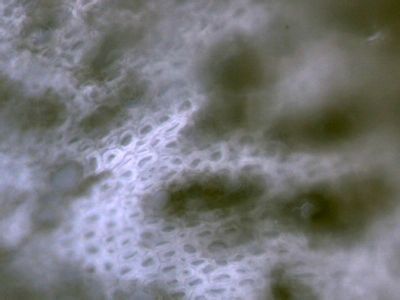
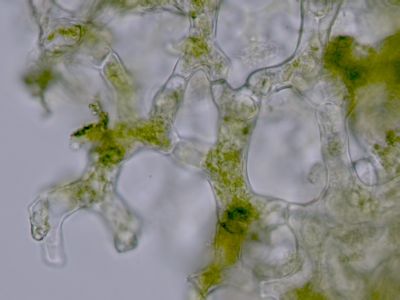
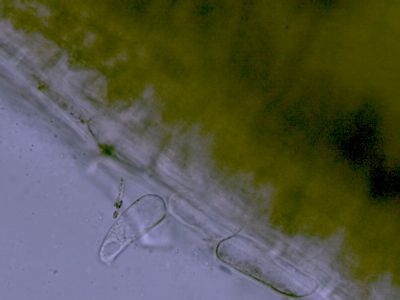
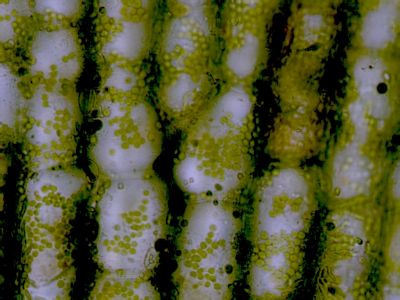
 Lístek mechu DicranumLístky mechů jsou tvořeny jednak parenchymatickými buňkami (1), jednak buňkami prosenchymatickými (2). Ty mají funkci mechanické opory lístku. © T.Č. Kučera 2006
Lístek mechu DicranumLístky mechů jsou tvořeny jednak parenchymatickými buňkami (1), jednak buňkami prosenchymatickými (2). Ty mají funkci mechanické opory lístku. © T.Č. Kučera 2006 -
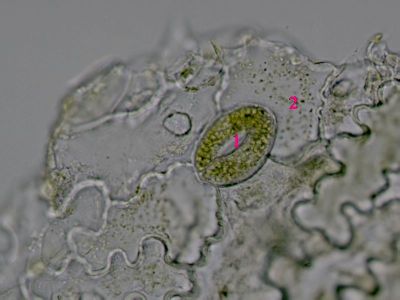
 Parenchymatická buňka z dužniny bobule Symphoricarpos albusDužnina bobulí pámelníku je tvořena parenchymatickými buňkami. Cytoplazma těchto buněk nevyplňuje buňku zcela, ale tvoří ramena (1), mezi nimiž jsou vakuoly (2). Nápadné je rovněž jádro (3). © T.Č. Kučera 2006
Parenchymatická buňka z dužniny bobule Symphoricarpos albusDužnina bobulí pámelníku je tvořena parenchymatickými buňkami. Cytoplazma těchto buněk nevyplňuje buňku zcela, ale tvoří ramena (1), mezi nimiž jsou vakuoly (2). Nápadné je rovněž jádro (3). © T.Č. Kučera 2006 -
 Parenchymatická buňka z dužniny bobule Symphoricarpos albus - 2Buňka dužniny bobule pámelníku s nápadným jádrem (1). Světlolomná struktura uvnitř jádra je jadérko. © T.Č. Kučera 2006
Parenchymatická buňka z dužniny bobule Symphoricarpos albus - 2Buňka dužniny bobule pámelníku s nápadným jádrem (1). Světlolomná struktura uvnitř jádra je jadérko. © T.Č. Kučera 2006 -
 Plastidy - chloroplastyDetail zelených plastidů - chloroplastů - v listech Elodea canadensis. © T.Č. Kučera 2006
Plastidy - chloroplastyDetail zelených plastidů - chloroplastů - v listech Elodea canadensis. © T.Č. Kučera 2006 -
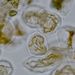 Plastidy - chromoplastyOranžověčervené plastidy, chromoplasty, představují typ fotosynteticky inaktivních plastidů. Jejich barva je dána přítomností karotenoidů. Detail parenchymatické buňky dužniny šípku. © T.Č. Kučera 2006
Plastidy - chromoplastyOranžověčervené plastidy, chromoplasty, představují typ fotosynteticky inaktivních plastidů. Jejich barva je dána přítomností karotenoidů. Detail parenchymatické buňky dužniny šípku. © T.Č. Kučera 2006 -
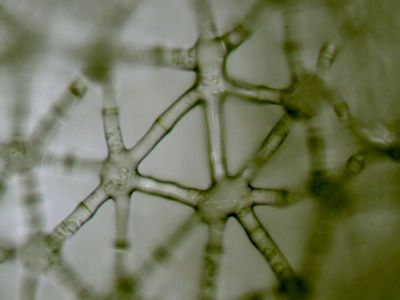
 Pletiva podle tvaru - aktinenchymAktinenchym je řazen mezi provzdušňovací pletiva. Je přítomen zejména ve stoncích bahnenních rostlin (zde z Juncus effusus). © T.Č. Kučera 2006
Pletiva podle tvaru - aktinenchymAktinenchym je řazen mezi provzdušňovací pletiva. Je přítomen zejména ve stoncích bahnenních rostlin (zde z Juncus effusus). © T.Č. Kučera 2006 -
 Pletiva podle tvaru - parenchymDřeňový parenchym stonku Pelargonium zonale. © T.Č. Kučera 2006
Pletiva podle tvaru - parenchymDřeňový parenchym stonku Pelargonium zonale. © T.Č. Kučera 2006 -
 Pletiva podle tvaru - rohový kolenchymKolenchymatická pletiva představují mechanickou oporu. Jsou tvořena buňkami s částečně ztlustlými stěnami. Na obrázku je rohový kolenchym, který má stěny ztlustlé na kontaktu tří a více buněk. Světlá místa představují stěny, tmavá lumen buněk. © T.Č. Kučera 2006
Pletiva podle tvaru - rohový kolenchymKolenchymatická pletiva představují mechanickou oporu. Jsou tvořena buňkami s částečně ztlustlými stěnami. Na obrázku je rohový kolenchym, který má stěny ztlustlé na kontaktu tří a více buněk. Světlá místa představují stěny, tmavá lumen buněk. © T.Č. Kučera 2006 -
 Pletiva podle tvaru - rohový kolenchymRohový kolenchym patří mez i mechanická pletiva. Stěny buněk jsou ztlustlé částečně, a to v rozích. Nachází obyvkle ve stoncích - zde v begónii (Begonia sp.). © T.Č. Kučera 2006
Pletiva podle tvaru - rohový kolenchymRohový kolenchym patří mez i mechanická pletiva. Stěny buněk jsou ztlustlé částečně, a to v rozích. Nachází obyvkle ve stoncích - zde v begónii (Begonia sp.). © T.Č. Kučera 2006 -
 Pletiva podle tvaru - sklerenchymSklerenchymtické buňky ve stonku. Nápadné jsou ztlustlé stěny buněk tohoto mechanického pletiva. © T.Č. Kučera 2006
Pletiva podle tvaru - sklerenchymSklerenchymtické buňky ve stonku. Nápadné jsou ztlustlé stěny buněk tohoto mechanického pletiva. © T.Č. Kučera 2006 -
 Plod - peckovice Prunus domesticaPlodem švestky je apokarpní peckovice. Perikarp peckovice je tvořen třemi vrtvami: blanitým exokarpem, dužnatým mezokarpem a sklerenchymatickým endokarpem (téý nazývaným sklerokarp). © T.Č. Kučera 2006
Plod - peckovice Prunus domesticaPlodem švestky je apokarpní peckovice. Perikarp peckovice je tvořen třemi vrtvami: blanitým exokarpem, dužnatým mezokarpem a sklerenchymatickým endokarpem (téý nazývaným sklerokarp). © T.Č. Kučera 2006 -
 Plod - šešule Alliaria petiolataPlodem čel. Brassicaceae mohou být tři různé plody: šešula, šešulka, struk a nažka. V případě česnáčku lékařského jde o šešuli - dvouplodolistový parakarpní plod, jehož podélný rozměr je větší, než příčný. V případě stejných nebo podobných rozměrů se plod označuje šešulka. Květy brukvovitých jsou čtyřčetné, heterochlamydní a bisymetrické. © T.Č. Kučera 2006
Plod - šešule Alliaria petiolataPlodem čel. Brassicaceae mohou být tři různé plody: šešula, šešulka, struk a nažka. V případě česnáčku lékařského jde o šešuli - dvouplodolistový parakarpní plod, jehož podélný rozměr je větší, než příčný. V případě stejných nebo podobných rozměrů se plod označuje šešulka. Květy brukvovitých jsou čtyřčetné, heterochlamydní a bisymetrické. © T.Č. Kučera 2006 -
 Plod - tobolky Hyoscyamus niger - detailPlodem blínu černého je tobolka, tedy plod, který se v čeledi Solanaceae vyskytuje méně často, než např. bobule. © T.Č. Kučera 2006
Plod - tobolky Hyoscyamus niger - detailPlodem blínu černého je tobolka, tedy plod, který se v čeledi Solanaceae vyskytuje méně často, než např. bobule. © T.Č. Kučera 2006 -
 Plod a větévka Prunus spinosaPlodem trnky obecné je, stejně jako u švestky a ostaních peckovin, apokarpní peckovice, jejíž perikarp má tři vrstvy oplodí. Květy a plody vyrůstají na zkrácených zašpičatělých větévkách - kolcích. © T.Č. Kučera 2006
Plod a větévka Prunus spinosaPlodem trnky obecné je, stejně jako u švestky a ostaních peckovin, apokarpní peckovice, jejíž perikarp má tři vrstvy oplodí. Květy a plody vyrůstají na zkrácených zašpičatělých větévkách - kolcích. © T.Č. Kučera 2006 -
 Plodenství - úborZ květenství vznikají po odkvětu plodenství. Plody u rodu Taraxacum sp. (ale i dalších zástupců čel. Asteraceae) jsou nažky (achenium) s chmýrem (pappus), který je kalichového původu a umožňuje anemochorii. © T.Č. Kučera 2006
Plodenství - úborZ květenství vznikají po odkvětu plodenství. Plody u rodu Taraxacum sp. (ale i dalších zástupců čel. Asteraceae) jsou nažky (achenium) s chmýrem (pappus), který je kalichového původu a umožňuje anemochorii. © T.Č. Kučera 2006 -
 Plodenství Carpinus betulusHroznovité plodenství habru obecného je složeno z oříšků, opatřených listenem. Ten představuje "křídlo" umožňující anemochorní šíření. © T.Č. Kučera 2006
Plodenství Carpinus betulusHroznovité plodenství habru obecného je složeno z oříšků, opatřených listenem. Ten představuje "křídlo" umožňující anemochorní šíření. © T.Č. Kučera 2006 -
 Plody - lusky Robinia pseudacaciaPlodem akátu je, stejně jako u většiny bobovitých, lusk. Jedná se o apokarpní jednoplodolistový pukavý plod. Lusky jsou u akátu uspořádány v hroznovité plodenství. © T.Č. Kučera 2006
Plody - lusky Robinia pseudacaciaPlodem akátu je, stejně jako u většiny bobovitých, lusk. Jedná se o apokarpní jednoplodolistový pukavý plod. Lusky jsou u akátu uspořádány v hroznovité plodenství. © T.Č. Kučera 2006 -
 Plody - tobolky Hyoscyamus nigerPlodem blínu černého je tobolka, tedy plod, který se v čeledi Solanaceae vyskytuje méně často, než např. bobule. © T.Č. Kučera 2006
Plody - tobolky Hyoscyamus nigerPlodem blínu černého je tobolka, tedy plod, který se v čeledi Solanaceae vyskytuje méně často, než např. bobule. © T.Č. Kučera 2006 -
 Podélný řez kořenem Ranunculus repensV záběru je patrná zejména rhizodermis (1). Objekt vpravo od podélného řezu (2) je pyramidální krystal šťavelanu vápenatého. © T.Č. Kučera 2006
Podélný řez kořenem Ranunculus repensV záběru je patrná zejména rhizodermis (1). Objekt vpravo od podélného řezu (2) je pyramidální krystal šťavelanu vápenatého. © T.Č. Kučera 2006 -
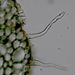 Příčný řez kořenem Ranuculus repens - detail kořenových vláskůKořenové vlásky několikanásobně zvětšují aktivní povrch kořene. Zakládají se v rhizodermis v tzv. trichoblastech. © T.Č. Kučera 2006
Příčný řez kořenem Ranuculus repens - detail kořenových vláskůKořenové vlásky několikanásobně zvětšují aktivní povrch kořene. Zakládají se v rhizodermis v tzv. trichoblastech. © T.Č. Kučera 2006 -
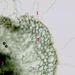 Příčný řez kořenem Ranunculus repensNa povrchu kořene je patrná jednovrstevná rhizodermis, z níž vyrůstají kořenové vlásky (1). Mohutná vrstva parenchymatických pletiv pod ní (2) představuje primární kůru. Tmavý útvar v levém dolním rohu řezu je střední válec. © T.Č. Kučera 2006
Příčný řez kořenem Ranunculus repensNa povrchu kořene je patrná jednovrstevná rhizodermis, z níž vyrůstají kořenové vlásky (1). Mohutná vrstva parenchymatických pletiv pod ní (2) představuje primární kůru. Tmavý útvar v levém dolním rohu řezu je střední válec. © T.Č. Kučera 2006 -
 Příčný řez kořenem Ranunculus repens - endodermis a střední válecNa příčném řezu je velice dobře patrná vnitřní vrstva primární kůry - endodermis (1) - se stlustlinami na antiklinách (=radiálně orientovaných stěnách) - tzv. Caspariho proužky. Směrem vpravo od endodermis následuje již střední válec, tvořený jednak vodivými pletivy (uspořádanými v případě kořene do radiálního cévního svazku) - 2 - a parenchymem (3). © T.Č. Kučera 2006
Příčný řez kořenem Ranunculus repens - endodermis a střední válecNa příčném řezu je velice dobře patrná vnitřní vrstva primární kůry - endodermis (1) - se stlustlinami na antiklinách (=radiálně orientovaných stěnách) - tzv. Caspariho proužky. Směrem vpravo od endodermis následuje již střední válec, tvořený jednak vodivými pletivy (uspořádanými v případě kořene do radiálního cévního svazku) - 2 - a parenchymem (3). © T.Č. Kučera 2006 -
 Příčný řez stonkem PelargoniumNa povrchu stonku je epidermis s kutinizovanou vnější periklinou (1). V primární kůře jsou přítomna mechanická pletiva - rohový kolenchym (2). Primární kůru od středního válce odděluje v blízkosti cévního svazku endodermis (3). Střední válec tvoří jednak základní pletiva (4), jednak cévní kolaterální svazky s částí lýkovou orientovanou centrifugálně (5) a částí dřevní orientovanou centripetálně (6). © T.Č. Kučera 2006
Příčný řez stonkem PelargoniumNa povrchu stonku je epidermis s kutinizovanou vnější periklinou (1). V primární kůře jsou přítomna mechanická pletiva - rohový kolenchym (2). Primární kůru od středního válce odděluje v blízkosti cévního svazku endodermis (3). Střední válec tvoří jednak základní pletiva (4), jednak cévní kolaterální svazky s částí lýkovou orientovanou centrifugálně (5) a částí dřevní orientovanou centripetálně (6). © T.Č. Kučera 2006 -
 Příčný řez stonkem begónie (Begonia sp.)Na řezu jsou patrné následující struktury: 1 - jednovrstevná epidermis s kutikulou, 2 - rohový kolenchym (součást primární kůry), 3 - asimilační pletivo stonku (chlorenchym) - rovněž součást primární kůry, 4 - parenchymatické základní pletivo středního válce. © T.Č. Kučera 2006
Příčný řez stonkem begónie (Begonia sp.)Na řezu jsou patrné následující struktury: 1 - jednovrstevná epidermis s kutikulou, 2 - rohový kolenchym (součást primární kůry), 3 - asimilační pletivo stonku (chlorenchym) - rovněž součást primární kůry, 4 - parenchymatické základní pletivo středního válce. © T.Č. Kučera 2006 -
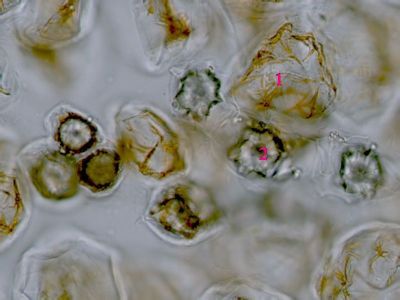
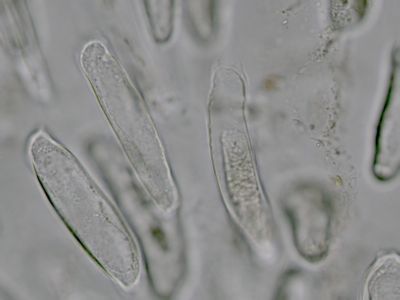
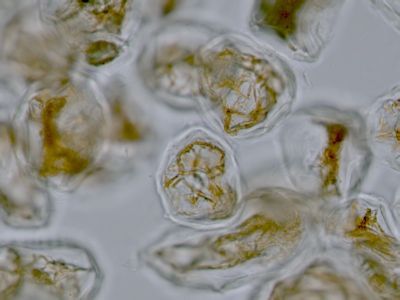
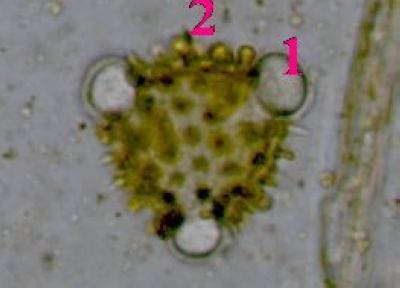
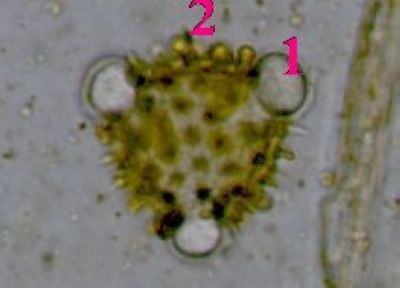
 Pylová zrna Senecio vulgarisPylová zrna krytosemenných rostlin představují mikrospory. Jejich obal (sporoderma) je tvořen dvěma vrstvami: vnitřní intinou a vnější exinou. V té jsou klíční póry (apertury) a výběžky umožňující přichycení na opylovače. © T.Č. Kučera 2006
Pylová zrna Senecio vulgarisPylová zrna krytosemenných rostlin představují mikrospory. Jejich obal (sporoderma) je tvořen dvěma vrstvami: vnitřní intinou a vnější exinou. V té jsou klíční póry (apertury) a výběžky umožňující přichycení na opylovače. © T.Č. Kučera 2006 -
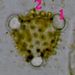 Pylové zrno Senecio vulgaris - detailToto pylové zrno má tři klíční póry (apertury)(1), jimiž proniká vnitřní vrstva sporodermy (intina). Ta je tvořena především pektiny, na rozdíl od vnější exiny, která je extrémně odolná díky impregnaci uhlovodíkem sporopoleninem. Výběžky (2) na povrchu exina usnadňují přichycení pylového zrna na tělo opylovače. © T.Č. Kučera 2006
Pylové zrno Senecio vulgaris - detailToto pylové zrno má tři klíční póry (apertury)(1), jimiž proniká vnitřní vrstva sporodermy (intina). Ta je tvořena především pektiny, na rozdíl od vnější exiny, která je extrémně odolná díky impregnaci uhlovodíkem sporopoleninem. Výběžky (2) na povrchu exina usnadňují přichycení pylového zrna na tělo opylovače. © T.Č. Kučera 2006 -
 Souplodí - Pulsatilla bohemicaSouplodí vznikají z květů s apokarpním gyneceem, které mají více jednoplodolistových pestíků. U konikleců (zde koniklece českého) je plodem nažka s chmýrem, usnadňujícím anemochorii. © T.Č. Kučera 2006
Souplodí - Pulsatilla bohemicaSouplodí vznikají z květů s apokarpním gyneceem, které mají více jednoplodolistových pestíků. U konikleců (zde koniklece českého) je plodem nažka s chmýrem, usnadňujícím anemochorii. © T.Č. Kučera 2006 -
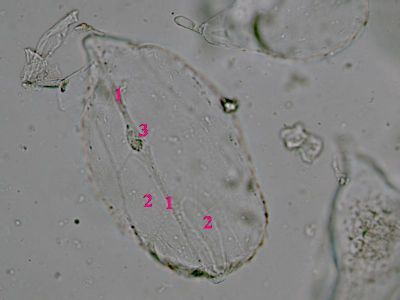
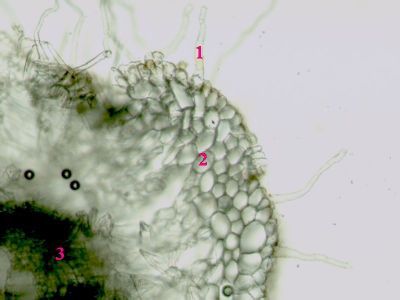
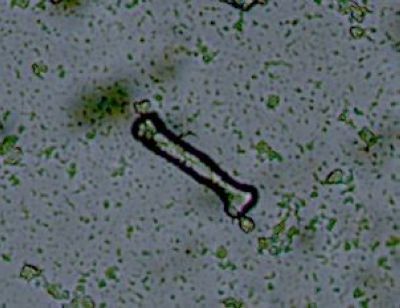
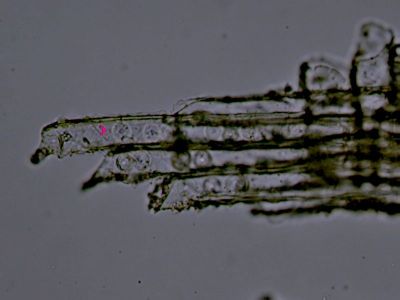
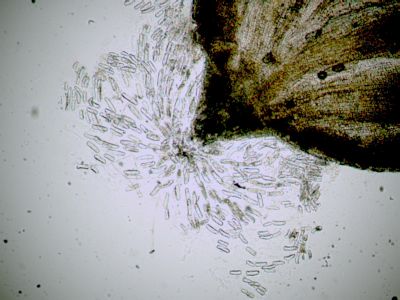
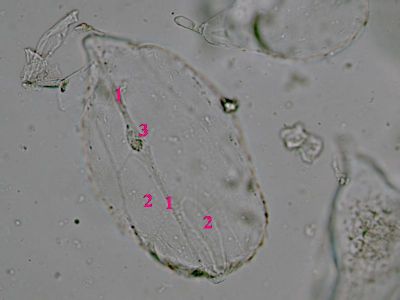
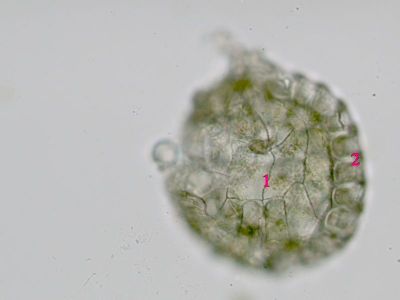
 Sporangium Polypodium vulgareDetailní pohled na sporangium osladiče. Jsou patrné buňky ve stěnách sporangia (1) a prstenec (annulus) (2), který umožňuje za sucha rozevření sporangia a uvolnění spor. © T.Č. Kučera 2006
Sporangium Polypodium vulgareDetailní pohled na sporangium osladiče. Jsou patrné buňky ve stěnách sporangia (1) a prstenec (annulus) (2), který umožňuje za sucha rozevření sporangia a uvolnění spor. © T.Č. Kučera 2006 -
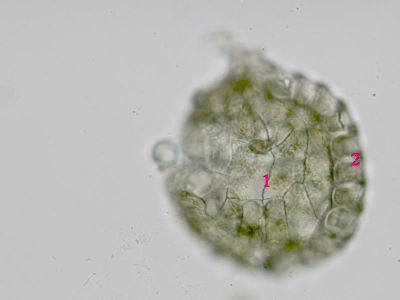
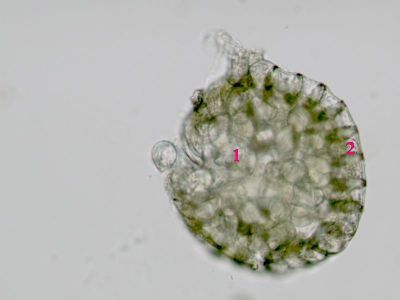
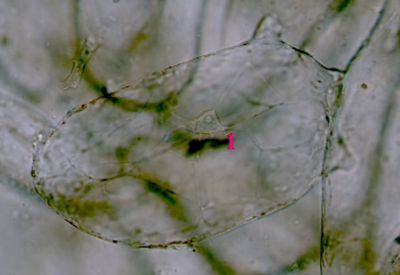
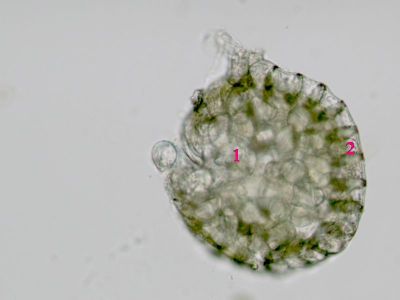
 Sporangium Polypodium vulgare - pohled do sporangiaUvnitř sporangia jsou patrné spory (1), v pravé části prstenec (annulus) - 2. © T.Č. Kučera 2006
Sporangium Polypodium vulgare - pohled do sporangiaUvnitř sporangia jsou patrné spory (1), v pravé části prstenec (annulus) - 2. © T.Č. Kučera 2006 -
 Stonek - brachyblasty Larix deciduaBrachyblasty představují větévky se zkrácenými internodii. Celá větévka je tak tvořena nahromaděnými nody. © T.Č. Kučera 2006
Stonek - brachyblasty Larix deciduaBrachyblasty představují větévky se zkrácenými internodii. Celá větévka je tak tvořena nahromaděnými nody. © T.Č. Kučera 2006 -
 Stonek - kolce Prunus spinosaKolce jsou, podobně jako brachyblasty, větévky se zkrácenými internodii. Na rozdíl od nich jsou zakončeny špičkou. Kolce jsou proto označovány jako trny stonkového původu. © T.Č. Kučera 2006
Stonek - kolce Prunus spinosaKolce jsou, podobně jako brachyblasty, větévky se zkrácenými internodii. Na rozdíl od nich jsou zakončeny špičkou. Kolce jsou proto označovány jako trny stonkového původu. © T.Č. Kučera 2006 -
 Stonek - normální větévky Larix deciduaKromě brachyblastů lze na modřínu opadavém nalézt i normální větévky s nezkrácenými internodii - jde o jednoleté výhonky, na nichž v dalším roce brachyblasty vyrůstají. Jehlice v tomto případě vyrůstají na větévce jednotlivě, ve spirále. © T.Č. Kučera 2006
Stonek - normální větévky Larix deciduaKromě brachyblastů lze na modřínu opadavém nalézt i normální větévky s nezkrácenými internodii - jde o jednoleté výhonky, na nichž v dalším roce brachyblasty vyrůstají. Jehlice v tomto případě vyrůstají na větévce jednotlivě, ve spirále. © T.Č. Kučera 2006 -
 Stonek - ovíjivý stonek Calystegia sepiumOpletník plotní má stejně jako většína ostatních zástupců čel. Convolvulaceae ovíjivý stonek. © T.Č. Kučera 2006
Stonek - ovíjivý stonek Calystegia sepiumOpletník plotní má stejně jako většína ostatních zástupců čel. Convolvulaceae ovíjivý stonek. © T.Č. Kučera 2006 -
 Stonek - příčný řezPohled na část příčného řezu stonkem. 1 - primární kůra, 2 - pericykl, 3 - parenchym středního válce, 4 - vodivá pletiva. © T.Č. Kučera 2006
Stonek - příčný řezPohled na část příčného řezu stonkem. 1 - primární kůra, 2 - pericykl, 3 - parenchym středního válce, 4 - vodivá pletiva. © T.Č. Kučera 2006 -
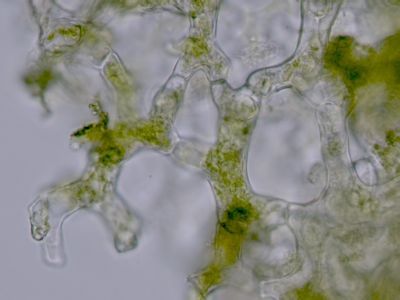
 Stonek - příčný řez 2Na povrchu stonku se zachovanou primární stavbou je zachována epidermis (1) se silnou kutikulou. © T.Č. Kučera 2006
Stonek - příčný řez 2Na povrchu stonku se zachovanou primární stavbou je zachována epidermis (1) se silnou kutikulou. © T.Č. Kučera 2006 -
 Stonek - sekundární stavba, příčný řez kmenem Robinia pseudacaciaNa řezu jsou patrné rozdíly mezi jádrovým dřevem (1) v nejstarší části sonku, které je impregnováno organickými látkami a bělovým dřevem (2) bez této impregnace. Sekundární lýko (3), deuterofloém, tvoří jen malou část hmoty kmene. Povrch je kmene je kryt sekundárními krycími pletivy, z nichž je nejnápadanější borka (rhytidoma). Praskliny ve vysychajícím dřevě obvykle kopírují směr radiálního parenchymu. © T.Č. Kučera 2006
Stonek - sekundární stavba, příčný řez kmenem Robinia pseudacaciaNa řezu jsou patrné rozdíly mezi jádrovým dřevem (1) v nejstarší části sonku, které je impregnováno organickými látkami a bělovým dřevem (2) bez této impregnace. Sekundární lýko (3), deuterofloém, tvoří jen malou část hmoty kmene. Povrch je kmene je kryt sekundárními krycími pletivy, z nichž je nejnápadanější borka (rhytidoma). Praskliny ve vysychajícím dřevě obvykle kopírují směr radiálního parenchymu. © T.Č. Kučera 2006 -
 Stonek - stvol Primula verisPrvosenky mají stvol, tj neolistěný květonosný stonek. Listy jsou v přízemní růžici. Vznik stvolu je vysvětlován nahloučením nodů v přízemní části, květonosný stonek představuje pak jediný článek stonku, nesoucí květenství (okolík). © T.Č. Kučera 2006
Stonek - stvol Primula verisPrvosenky mají stvol, tj neolistěný květonosný stonek. Listy jsou v přízemní růžici. Vznik stvolu je vysvětlován nahloučením nodů v přízemní části, květonosný stonek představuje pak jediný článek stonku, nesoucí květenství (okolík). © T.Č. Kučera 2006 -
 Trichocereus atacamensisCactoideae: Velký kaktus, který může dorůstat až do 15 m výšky. Vyskytuje se v suchých kamenitých oblastech Bolivie a v severní části Argentiny, kde stoupá až do 3500 m n.m. Valle Fertil, Argentina. © Michal Mergl, 2015
Trichocereus atacamensisCactoideae: Velký kaktus, který může dorůstat až do 15 m výšky. Vyskytuje se v suchých kamenitých oblastech Bolivie a v severní části Argentiny, kde stoupá až do 3500 m n.m. Valle Fertil, Argentina. © Michal Mergl, 2015 -
 Trichocereus atacamensis
Trichocereus atacamensis
kaktusCactoideae Velký kaktus, který m?že dor?stat až do 15 m výšky. Vyskytuje se v suchých kamenitých oblastech Bolivie a v severní ?ásti Argentiny, kde stoupá až do 3500 m n.m. Valle Fertil, Argentina. © Michal Mergl, 2015 -
 Trichomy - krycí trichomTrichomy jsou deriváty epidermis, zakládají se jako trichoblasty. V tomto případě jde o krycí trichom zakončený špičkou. Pelargonium zonale. © T.Č. Kučera 2006
Trichomy - krycí trichomTrichomy jsou deriváty epidermis, zakládají se jako trichoblasty. V tomto případě jde o krycí trichom zakončený špičkou. Pelargonium zonale. © T.Č. Kučera 2006 -
 Trny palistového původuTrny trnovníku akátu (Robinia pseudacacia) jsou palistového původu. Dokládá to jednak jejich párovitost, jednak růst na bázi řapíku. © T.Č. Kučera 2006
Trny palistového původuTrny trnovníku akátu (Robinia pseudacacia) jsou palistového původu. Dokládá to jednak jejich párovitost, jednak růst na bázi řapíku. © T.Č. Kučera 2006 -
 Trny palistového původuTrny trnovníku akátu (Robinia pseudacacia) jsou palistového původu. Dokládá to jednak jejich párovitost, jednak růst na bázi řapíku. Nekrotické skvrny na listech jou způsobeny parizitickou klíněnkou akátovou (Phyllonorycter robiniella). © T.Č. Kučera 2006
Trny palistového původuTrny trnovníku akátu (Robinia pseudacacia) jsou palistového původu. Dokládá to jednak jejich párovitost, jednak růst na bázi řapíku. Nekrotické skvrny na listech jou způsobeny parizitickou klíněnkou akátovou (Phyllonorycter robiniella). © T.Č. Kučera 2006 -
 Úbor Taraxacum sp.Květenstvím čeledi Asteraceae s. lato je úbor. V případě pamapelišky, která patří do čeledi Cichoriaceae, je úbor tvořen pouze jazykovitými květy, trubkovité se nevyskytují. Na bázi úboru je zákrov listenového původu (na foto není patrný). © T.Č. Kučera 2006
Úbor Taraxacum sp.Květenstvím čeledi Asteraceae s. lato je úbor. V případě pamapelišky, která patří do čeledi Cichoriaceae, je úbor tvořen pouze jazykovitými květy, trubkovité se nevyskytují. Na bázi úboru je zákrov listenového původu (na foto není patrný). © T.Č. Kučera 2006 -
 Vodivá pletiva - cévní svazek podélněKolaterální cévní svazek. Dřevní část svazku je tvořena primárním dřevem: 1 - metaxylém, 2 - protoxylém; 3 - krystal šťavelanu vápenatého. © T.Č. Kučera 2006
Vodivá pletiva - cévní svazek podélněKolaterální cévní svazek. Dřevní část svazku je tvořena primárním dřevem: 1 - metaxylém, 2 - protoxylém; 3 - krystal šťavelanu vápenatého. © T.Č. Kučera 2006 -
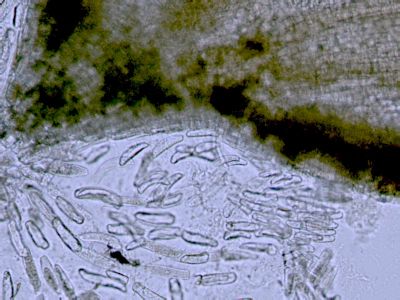
 Vodivá pletiva - pohled do cévního svazkuPodélný řez cévním svazkem. 1 - spirálně a žebříčkovitě ztlustlé stěny jsou charakteristické pro cévy primárního xylému. 2 - Sítkovice jsou naproti tomu tenkostěnné trubicovité struktury, které mají zachovány příčné přepážky. © T.Č. Kučera 2006
Vodivá pletiva - pohled do cévního svazkuPodélný řez cévním svazkem. 1 - spirálně a žebříčkovitě ztlustlé stěny jsou charakteristické pro cévy primárního xylému. 2 - Sítkovice jsou naproti tomu tenkostěnné trubicovité struktury, které mají zachovány příčné přepážky. © T.Č. Kučera 2006 -
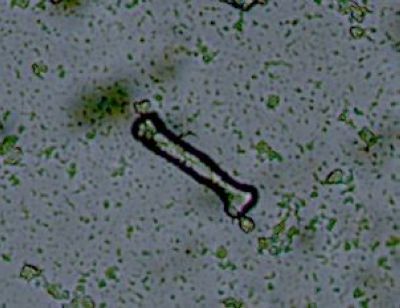
 Zásobní látky - hnátovitý typ škrobových zrnHnátovitý typ škrobových zrn připomínající tvarem dlouhé kosti, je charakteristický pro pryšcovité rostliny (Euphorbiaceae). Latex z Euphorbia leuconeura. © T.Č. Kučera 2006
Zásobní látky - hnátovitý typ škrobových zrnHnátovitý typ škrobových zrn připomínající tvarem dlouhé kosti, je charakteristický pro pryšcovité rostliny (Euphorbiaceae). Latex z Euphorbia leuconeura. © T.Č. Kučera 2006





































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2016 Tomáš Jakeš a Jan Hodinář